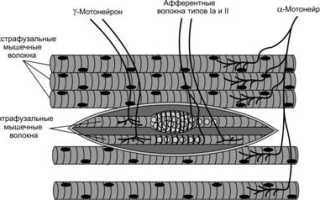
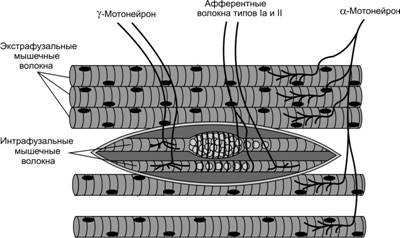
Существует два основных типа специфических рецепторов в скелетной мышце: мышечные веретена и рецепторы Гольджи (сухожильные рецепторы). В этой статье рассматриваются строение и функции мышечных веретен. Описаны также интрафузальные мышечные волокна, которые входят в состав этих веретен.
РЕЦЕПТОРЫ СКЕЛЕТНОЙ МЫШЦЫ (МЫШЕЧНЫЕ ВЕРЕТЕНА)
Познакомившись со строением нервов, можно обсудить рецепторы скелетной мышцы. Выделяют два основных типа рецепторов: мышечные веретена и рецепторы Гольджи (сухожильные рецепторы Гольджи).
Мышечные веретена были описаны в середине XIX века немецким физиологом Вильгельмом Кюне (Wilhelm Kühne, 1863). Они представляют собой вытянутые структуры, расширенные посередине, напоминающие веретено, использовавшееся в прядении. Английский физиолог Чарльз Скотт Шеррингтон первым указал на чувствительность мышечных веретен.
Мышечные веретена расположены внутри мышцы параллельно мышечным волокнам и прикрепляются к эндомизию, окружающему волокна, или к сухожилию. Длина веретена достигает 10 мм, а ширина в самой широкой части — от 80 до 250 мкм (рис. 1).
Рис. 1. Строение мышечного веретена
Количество мышечных веретен в мышце варьирует от десятков до сотен. Например, в двуглавой мышце плеча — 320 веретен, в большой грудной — 450, а в трехглавой — 520. Плотность веретен (количество на 1 г массы мышцы) также различна: наибольшая в тыльных мышцах шеи, наименьшая — в мышцах конечностей. В верхней косой мышце головы плотность составляет 42,7 на 1 г, а в двуглавой — 2,0 на 1 г.
Мышечные веретена возбуждаются при растяжении мышцы, что происходит, например, во время упражнений на растяжение или эксцентрических нагрузок. Растяжение также возникает при ходьбе, беге, прыжках или езде на велосипеде.
Информация от рецепторов мышечных веретен сообщает нервной системе текущую длину мышцы и скорость ее растяжения.
Внутри мышечного веретена находятся тонкие интрафузальные мышечные волокна, диаметр которых в 2-3 раза меньше обычных экстрафузальных. Интрафузальные волокна могут сокращаться и расслабляться, их количество в веретене составляет от 4 до 14. Эти волокна имеют собственную иннервацию, что позволяет регулировать чувствительность веретена. К ним подходят двигательные мотонейроны (γ-мотонейроны), от которых отходят чувствительные волокна, передающие информацию о длине и скорости растяжения мышцы в ЦНС. За открытия в этой области шведскому физиологу Рагнару Граниту была присуждена Нобелевская премия.
Информация от мышечных веретен помогает предотвращать повреждения мышцы и служит основой для функционирования рефлекса на растяжение.
 Читайте также:
Читайте также:

Литература
- Гранит Р. Основы регуляции движений. — М.: Мир, 1973. — 278 с. Ил.
- Мак-Комас, А.Дж. Скелетные мышцы. — Киев: Олимпийская литература, 2001. — 407 с.
- Самсонова А.В. Моторные и сенсорные компоненты биомеханической структуры физических упражнений: автореф. дис. … докт. пед. наук. — СПб, 1998. — 48 с.
- Самсонова А.В., Комиссарова Е.Н. Биомеханика мышц: учебно-методическое пособие / Под ред. А.В. Самсоновой. — Санкт-Петербургский гос. ун-т физической культуры им. П.Ф. Лесгафта, 2008. — 127 с.
- Самсонова А.В. Гипертрофия скелетных мышц человека. — СПб: Кинетика, 2018. — 159 с.
С уважением, А.В. Самсонова
Похожие записи:
Томас ДеЛорме — ученый, разработавший метод прогрессивно возрастающего сопротивления
Врач-реабилитолог Томас ДеЛорме разработал специальные силовые тренажеры для своих пациентов и использовал большие отягощения для улучшения их состояния.
Мышечные боли или почему болят мышцы после тренировок? (запаздывающие болезненные ощущения)
Рассмотрены виды и причины болей в мышцах, возникающих через день после тренировки. Объясняется, что основная причина мышечных болей…
Мышечные боли или почему болят мышцы во время и после тренировок? (острые болезненные ощущения)
Изучены виды и причины болей в мышцах после тренировки, с акцентом на боли, возникающие во время…
От чего зависит сила мышц? (физиологические факторы)
Обсуждаются физиологические факторы, определяющие силу скелетных мышц человека: частота импульсации двигательных единиц, количество активных единиц, синхронизация…
Классификация соматотипов детей по Штефко-Островскому
Представлена классификация типов телосложения детей по Штефко-Островскому: астеноидный, торакальный, мышечный и дигестивный. Кратко изложен жизненный путь В.Г. Штефко.
Типы телосложения (соматотипы) по Хит-Картеру
Рассмотрены типы телосложения (соматотипы) по классификации Б.Х. Хит и Д.Э.Л. Картера. Описаны преимущества данной классификации по сравнению с…
Мышечное веретено — Muscle spindle
Иннервируемая мышечная структура, участвующая в рефлекторных действиях и проприоцепции
- Читайте также:
| Мышечное веретено | |
|---|---|
| Мышечное веретено млекопитающих: типичное положение в мышце (слева), нейронные связи в спинном мозге (в центре) и расширенная схема (справа). Веретено — это рецептор растяжения с собственным двигателем, состоящий из нескольких интрафузальных мышечных волокон. Сенсорные окончания первичной афферентной группы (Ia) и вторичной группы (II) обвивают неконтрактильные центральные части интрафузальных волокон. Гамма-мотонейроны активируют интрафузальные волокна, изменяя скорость возбуждения в состоянии покоя и чувствительность афферентов к растяжению. | |
| Подробности | |
| Часть | мышечной системы |
| Скелетно-мышечный | |
| Идентификаторы | |
| Латинский | fusus neromuscularis |
| MeSH | D009470 |
| TH | H3.11.06.0.00018 |
| FMA | 83607 |
| Анатомическая терминология [правка в Викиданных ] |
Мышечные веретена — это рецепторы растяжения внутри мышцы, которые обнаруживают изменения её длины. Они передают информацию о длине в центральную нервную систему через афферентные нервные волокна. Эта информация обрабатывается мозгом как проприоцепция. Ответы мышечных веретен на изменение длины играют важную роль в регулировании сокращения мышц, например, активируя мотонейроны через рефлекс растяжения, чтобы противостоять растяжению.
Мышечное веретено имеет как сенсорные, так и моторные компоненты:
- Сенсорная информация передается первичными сенсорными волокнами типа Ia, которые спирально обвивают мышечные волокна внутри веретена, и вторичными сенсорными волокнами типа II.
- Активация мышечных волокон внутри веретена осуществляется гамма-мотонейронами и в меньшей степени бета-мотонейронами.
Структура
Мышечные веретена расположены внутри мышц, между экстрафузальными мышечными волокнами. Специализированные волокна, составляющие мышечное веретено, называются интрафузальными волокнами, в отличие от экстрафузальных волокон, которые образуют основную массу мышцы. Мышечные веретена имеют капсулу из соединительной ткани и располагаются параллельно экстрафузальным волокнам.
Состав
Мышечные веретена состоят из 5-14 мышечных волокон, которые делятся на три типа: динамические волокна ядерного мешка (мешок 1), статические волокна ядерного мешка (мешок 2) и волокна ядерной цепи.
Первичные сенсорные волокна типа Ia (большого диаметра) обвивают интрафузионные мышечные волокна, заканчиваясь около середины каждого из них. Вторичные сенсорные волокна типа II (среднего диаметра) заканчиваются в центральных областях статического мешка и волокон цепи. Эти волокна передают информацию через механически управляемые ионные каналы аксонов, чувствительных к растяжению.
Моторная часть веретена представлена моторными нейронами: до дюжины гамма-мотонейронов и один или два бета-мотонейрона, которые вместе называются фузимоторными нейронами. Они активируют мышечные волокна веретена. Гамма-двигательные нейроны иннервируют только волокна внутри веретена, тогда как бета-двигательные нейроны — как внутри, так и снаружи. Активация нейронов вызывает сокращение и жесткость концов мышечных волокон веретена.
Фузимоторные нейроны делятся на статические и динамические в зависимости от типа иннервируемых мышечных волокон и их влияния на ответы сенсорных нейронов Ia и II, которые иннервируют центральную неконтрактильную часть веретена.
- Статические аксоны иннервируют цепочку или волокна статического мешка 2. Они увеличивают скорость возбуждения афферентов Ia и II при фиксированной длине мышцы.
- Динамические аксоны иннервируют мешок 1 интрафузионных мышечных волокон, повышая чувствительность афферентов Ia к растяжению.
Эфферентные нервные волокна гамма-мотонейронов также заканчиваются в мышечных веретенах, образуя синапсы на концах интрафузальных волокон и регулируя чувствительность сенсорных афферентов в неконтрактильной центральной области.
Функция
Рефлекс растяжения
Когда мышца растягивается, первичные сенсорные волокна мышечного веретена типа Ia реагируют на изменение длины и скорость растяжения, передавая эту информацию в спинной мозг в виде изменений частоты потенциалов действия. Вторичные сенсорные волокна типа II также реагируют на изменения длины мышцы, но с меньшей чувствительностью к скорости, и передают сигнал в спинной мозг. Афферентные сигналы Ia моносинаптически передаются множеству альфа-мотонейронов мышцы, содержащей рецепторы. Рефлекторная активность альфа-мотонейронов передается через их эфферентные аксоны к экстрафузальным волокнам мышцы, которые генерируют силу и сопротивляются растяжению. Афферентный сигнал Ia также передается полисинаптически через интернейроны (ингибирующие интернейроны Ia), которые ингибируют альфа-мотонейроны антагонистических мышц, вызывая их расслабление.
- Читайте также:
Модификация чувствительности
Функция гамма-мотонейронов заключается не в увеличении силы сокращения мышц, а в изменении чувствительности сенсорных афферентов мышечного веретена к растяжению. При высвобождении ацетилхолина гамма-мотонейроном сокращаются концевые части интрафузальных мышечных волокон, что удлиняет неконтрактильные центральные части (см. схему «фузимоторного действия» ниже). Это открывает ионные каналы, чувствительные к растяжению, в сенсорных окончаниях, что приводит к притоку ионов натрия. В результате повышается потенциал покоя окончаний, увеличивая вероятность срабатывания потенциала действия и, соответственно, чувствительность афферентов мышечного веретена к растяжению.
Как центральная нервная система контролирует гамма-фузимоторные нейроны? Запись гамма-мотонейронов во время нормального движения затруднена из-за их маленьких аксонов. Предложено несколько теорий на основе записей афферентов веретена:
- Коактивация альфа-гамма. Гамма-мотонейроны активируются параллельно с альфа-мотонейронами, чтобы поддерживать активацию афферентов веретена при укорачивании экстрафузальных мышц.
- Фузимоторный набор. Гамма-мотонейроны активируются в зависимости от новизны или сложности задачи. Статические гамма-мотонейроны постоянно активны во время рутинных движений, таких как локомоция, в то время как динамические гамма-мотонейроны активизируются чаще при сложных задачах, повышая чувствительность к растяжению Ia.
- Фузимоторный шаблон предполагаемого движения. Статическая гамма-активность представляет собой «временной шаблон» ожидаемого укорачивания и удлинения мышцы. Динамическая гамма-активность внезапно включается и выключается, повышая чувствительность афферентов веретена к началу удлинения мышц и отклонениям от намеченной траектории движения.
Развитие
Считается, что веретена играют ключевую роль в сенсомоторном развитии.
Клиническое значение
После инсульта или повреждения спинного мозга у человека может развиться спастическая гипертония (спастический паралич). Это состояние приводит к чрезмерной чувствительности рефлекса растяжения в мышцах-сгибателях рук и мышцах-разгибателях ног. В результате возникают неправильная поза, скованность и контрактуры. Гипертония может быть следствием повышенной чувствительности альфа-мотонейронов и интернейронов к афферентным сигналам Ia и II.
Дополнительные изображения
- Мышечное веретено
- Гамма-волокно
- 1A-волокно
- Альфа-волокно
- Схема фузимоторного действия
См. Также
- Сенсорное волокно типа Ia
- Сенсорное волокно типа II
- Альфа-мотонейрон
- Гамма-мотонейрон
- Бета-мотонейрон
- Интрафузальное мышечное волокно
- Экстрафузальные мышечные волокна
Примечания
Ссылки
Внешние ссылки
- Мышцы и веретена в Медицинских предметных рубриках Национальной медицинской библиотеки США (MeSH)
Структура и функции проприорецепторов
В активной части опорно-двигательного аппарата находятся проприорецепторы, которые делятся на два типа: нервно-мышечные веретёна, реагирующие на изменение длины мышцы, и сухожильные органы Гольджи, чувствительные к изменению напряжения (Рисунок 1).
Нервно-мышечные веретёна расположены в скелетных мышцах между мышечными волокнами. Они представляют собой веретеновидные структуры, покрытые соединительнотканной капсулой. Длина веретена составляет 4–7 мм (до 10 мм), диаметр — 100 мкм. Внутри веретена находятся видоизменённые мышечные волокна — интрафузальные, которые отличаются от обычных экстрафузальных. Интрафузальные волокна имеют меньший диаметр (15–30 мкм) и не содержат сократительных элементов в центральной части, что позволяет сокращаться только периферическим участкам. Концы интрафузального волокна фиксированы к капсуле, и при укорочении периферических отделов центральная часть растягивается. Интрафузальные волокна располагаются параллельно экстрафузальным и делятся на два типа: с ядерной сумкой и с ядерной цепочкой (Рисунок 2). Волокна с ядерной сумкой подразделяются на статические и динамические. Первые имеют расширенную центральную часть с собранными в скопления ядрами, вторые — тоньше и с ядрами, расположенными в ряд. В каждом веретене содержится 1–3 волокна с ядерной сумкой и 5–7 с ядерной цепочкой.
Интрафузальные волокна получают афферентную и эфферентную иннервацию. Выделяют два типа афферентных окончаний: первичные (аннулоспиральные) и вторичные. Первичные окончания обвивают центральную часть интрафузальных волокон и иннервируются волокнами типа Ia, которые являются самыми толстыми (около 17 мкм) и имеют высокую скорость проведения (70–120 м/с). Каждое веретено иннервируется одним волокном Ia, которое получает импульсацию от всех первичных окончаний. Вторичные окончания имеют спиралевидную форму и оплетают центральные части волокон с ядерной цепочкой и статических волокон с ядерной сумкой, но отсутствуют у динамических волокон. Они иннервируются волокнами типа II (около 8 мкм) со скоростью 40–70 м/с (Рисунок 3).
Оба типа окончаний чувствительны к изменению длины интрафузальных волокон, но реагируют по-разному. При медленном растяжении импульсация возникает в обоих типах окончаний, а при быстром — только в первичных, что приводит к динамическому ответу. Волокна Ia передают информацию о длине и скорости изменения длины, тогда как волокна II — только о длине.
Эфферентная иннервация представлена волокнами типа Aγ, которые являются аксонами γ-мотонейронов спинного мозга. Эти волокна иннервируют периферические отделы интрафузальных волокон, вызывая их сокращение и растяжение центральной части. Выделяют γ-динамические и γ-статические волокна, которые повышают чувствительность рецепторов.
-
Читайте также:
При увеличении длины мышцы рецепторы нервно-мышечных веретён возбуждаются, и импульсация по волокнам Ia и II поступает в спинной мозг. Здесь афферентные нейроны образуют синапсы с α-моторными нейронами, вызывая сокращение экстрафузальных волокон и укорочение мышцы. Одновременно коллатерали афферентных нейронов активируют тормозные вставочные нейроны, угнетающие α-мотонейроны антагонистов. Этот рефлекс называется миотатическим и имеет небольшой латентный период (~0,9 мс).
Различают фазический (динамический) и тонический миотатический рефлекс. Фазический рефлекс активируется при быстром растяжении, а тонический — при медленном, поддерживая вертикальную позу. Например, при утомлении у человека в стойке «смирно» мышцы растягиваются, и миотатический рефлекс возвращает тело в правильное положение.
При возбуждении γ-мотонейронов сокращаются периферические отделы интрафузальных волокон, что растягивает центральную часть. Это активирует первичные и вторичные окончания, запуская миотатический рефлекс. Такой механизм называется γ-петлёй и обеспечивает сохранение чувствительности рецепторов при изменении длины мышцы. α-γ-коактивация позволяет сохранять чувствительность рецепторов как при сокращении, так и при расслаблении мышцы (Рисунок 6).
Сухожильные органы Гольджи представляют собой сеть коллагеновых нитей, окружённых соединительнотканной капсулой. Они расположены в месте прикрепления мышечных волокон к сухожилиям и реагируют на напряжение, а не на длину мышцы. Афферентные волокна от сухожильных органов Гольджи следуют в спинной мозг и переключаются на вставочные тормозные нейроны, что приводит к расслаблению мышцы. Этот рефлекс называется обратным миотатическим рефлексом и предотвращает чрезмерное напряжение мышц, а также обеспечивает равномерность силы сокращения различных волокон.
Импульсация от проприорецепторов поступает не только в спинной мозг, но и в различные отделы головного мозга, включая мозжечок и кору больших полушарий. Эта информация необходима для регуляции движений и осознания положения частей тела в пространстве (Рисунок 8).
- Wise, «Review of the History of Medicine» (Л., 1967).
- Мустафин Р. И. и др. «Сравнительное исследование поликомплексных систем для гастроретентивной доставки метформина». Разработка и регистрация лекарственных средств. 2015; 1(10): 48–50.
- https://allasamsonova.ru/receptory-skeletnoj-myshcy-myshechnye-veretena/.
- https://wikichi.ru/wiki/Muscle_spindle.
- https://manumed.pro/%D1%81%D1%82%D1%80%D1%83%D0%BA%D1%82%D1%83%D1%80%D0%B0-%D0%B8-%D1%84%D1%83%D0%BD%D0%BA%D1%86%D0%B8%D0%B8-%D0%BF%D1%80%D0%BE%D0%BF%D1%80%D0%B8%D0%BE%D1%80%D0%B5%D1%86%D0%B5%D0%BF%D1%82%D0%BE%D1%80/.
- Haeser, «Handbuch der Gesch. d. Medicin».