А-ДНК – сухая молекула
А-форма — это правый винт с 11 комплементарными парами оснований в каждом витке. Диаметр составляет 2,3 нм, а длина одного витка спирали — 2,5 нм. Плоскости, образуемые спаренными основаниями, наклонены на 20° к оси молекулы. Соседние нуклеотиды расположены компактно, на расстоянии 0,23 нм друг от друга.
Эта форма ДНК возникает при низкой гидратации и высокой концентрации ионов натрия и калия. Она характерна для процессов, в которых ДНК образует комплекс с РНК, так как РНК не может принимать другие формы. А-форма также устойчива к ультрафиолетовому облучению и встречается в дезоксирибонуклеиновой кислоте грибных спор.
Суперскрученная молекула
Сверхспирализованная форма ДНК возникает, когда двухцепочечная молекула дополнительно закручивается. Закручивание может быть положительным (по часовой стрелке) или отрицательным (против часовой стрелки). У большинства организмов ДНК суперскручена отрицательно, то есть против основных витков двойной спирали.
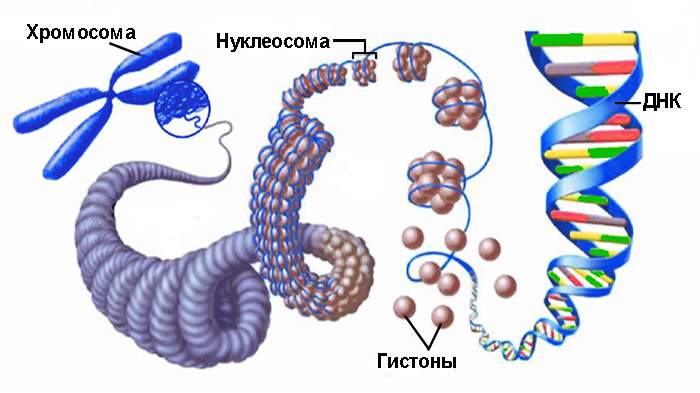
При образовании дополнительных петель — супервитков — ДНК принимает сложную пространственную конфигурацию. В клетках эукариот этот процесс включает формирование комплексов, в которых ДНК отрицательно навивается на гистоновые белки, образуя нити с нуклеосомами, напоминающими бусины. Свободные участки нити называются линкерами. В поддержании суперскрученной формы молекулы ДНК участвуют также негистоновые белки и неорганические соединения, что приводит к образованию хроматина — вещества хромосом.
Хроматиновые нити с нуклеосомами могут усложнять свою морфологию в процессе, называемом конденсацией хроматина.
Этапы синтеза
Все процессы матричного синтеза делятся на три этапа:
- инициация;
- элонгация;
- терминация.
Инициация — это подготовка к синтезу, которая зависит от типа процесса. Основная цель этой стадии — привести систему фермент-субстрат в рабочее состояние.
Во время элонгации происходит наращивание синтезируемой цепи. Между звеньями, подобранными согласно матричной последовательности, образуется ковалентная связь (пептидная или фосфодиэфирная). Терминация завершает синтез и освобождает продукт.
Строение и функции ДНК
ДНК — полимер, мономерами которого являются дезоксирибонуклеотиды. Модель двойной спирали молекулы ДНК была предложена в 1953 году Дж. Уотсоном и Ф. Криком, основываясь на работах М. Уилкинса, Р. Франклин и Э. Чаргаффа.
- Читайте также:
Молекула ДНК состоит из двух полинуклеотидных цепей, закрученных спирально друг вокруг друга и вокруг воображаемой оси. Исключение составляют некоторые вирусы с одноцепочечной ДНК. Диаметр двойной спирали составляет 2 нм, расстояние между соседними нуклеотидами — 0,34 нм, а на один оборот спирали приходится 10 пар нуклеотидов. Длина молекулы может достигать нескольких сантиметров, а молекулярный вес — десятков и сотен миллионов. Суммарная длина ДНК в ядре клетки человека составляет около 2 м. В эукариотических клетках ДНК образует комплексы с белками и имеет специфическую пространственную конформацию.
Мономер ДНК — дезоксирибонуклеотид — состоит из трех компонентов: 1) азотистого основания, 2) пятиуглеродного моносахарида (дезоксирибозы) и 3) фосфорной кислоты. Азотистые основания делятся на пиримидины и пурины. Пиримидиновые основания ДНК — тимин и цитозин, а пуриновые — аденин и гуанин.
Название нуклеотида происходит от названия соответствующего основания. Нуклеотиды и азотистые основания обозначаются заглавными буквами.
| Азотистое основание | Название нуклеотида | Обозначение |
|---|---|---|
| Аденин | Адениловый | А (A) |
| Гуанин | Гуаниловый | Г (G) |
| Тимин | Тимидиловый | Т (T) |
| Цитозин | Цитидиловый | Ц (C) |
Полинуклеотидная цепь образуется в результате реакций конденсации нуклеотидов. При этом между 3′-углеродом дезоксирибозы одного нуклеотида и остатком фосфорной кислоты другого образуется фосфоэфирная связь. Один конец цепи называется 5′-концом, а другой — 3′-концом.
Против одной цепи располагается вторая. Расположение нуклеотидов в этих цепях строго определено: аденин всегда соединяется с тимином, а гуанин — с цитозином. Между аденином и тимином образуются две водородные связи, между гуанином и цитозином — три. Эта закономерность называется принципом комплементарности. Дж. Уотсон и Ф. Крик пришли к пониманию этого принципа, основываясь на работах Э. Чаргаффа, который установил, что в любом фрагменте ДНК содержание гуанина соответствует содержанию цитозина, а аденина — тимину («правило Чаргаффа»).
Из принципа комплементарности следует, что последовательность нуклеотидов одной цепи определяет последовательность нуклеотидов другой.
Цепи ДНК антипараллельны, то есть нуклеотиды располагаются в противоположных направлениях: напротив 3′-конца одной цепи находится 5′-конец другой. Молекулу ДНК иногда сравнивают с винтовой лестницей, где «перила» — это сахарофосфатный остов, а «ступени» — комплементарные азотистые основания.
Основная функция ДНК — хранение и передача наследственной информации.
Нуклеиновые кислоты – полимерные молекулы
Нуклеиновые кислоты — это крупные полимерные органические молекулы, известные как полинуклеотиды. Обычно ДНК значительно больше РНК. Их мономерами являются нуклеотиды, состоящие из трех компонентов:

- пентозы (рибоза в РНК и дезоксирибоза в ДНК);
- фосфатной группы (остаток фосфорной кислоты —PO₄);
- азотистого основания.
Азотистые основания — это ароматические гетероциклические соединения, производные пиримидина или пурина. Существует пять основных типов азотистых оснований. Двухкольцевые пурины: аденин (A) и гуанин (G) встречаются как в ДНК, так и в РНК. Остальные три основания — однокольцевые молекулы, производные пиримидина: цитозин (C — есть в ДНК и РНК), тимин (T — только в ДНК) и урацил (U — только в РНК).
Аденин и рибоза образуют нуклеозид аденозин (A). Производные других азотистых оснований называются гуанозин (G), уридин (U), тимидин (T) и цитидин (C). При соединении азотистых оснований с дезоксирибозой образуются дезоксинуклеозиды. Все нуклеозидфосфаты объединяют под общим названием — нуклеотиды.
Нуклеиновые кислоты формируются в результате реакции обезвоживания (конденсации) между фосфатной группой одного нуклеотида и гидроксильной группой пентозы другого. Это приводит к образованию фосфодиэфирной связи, соединяющей два углевода через фосфат.
В молекуле нуклеотида азотистое основание присоединено к первому атому углерода пентозы, а остаток фосфорной кислоты — к пятому. Полинуклеотидная цепь полярна и имеет два конца:
- 5′ (пять-штрих положение) — углеродный атом в рибозе или дезоксирибозе;
- 3′ (три-штрих положение) — гидроксильная группа от углевода (ОН).
В двойной спирали ДНК эти концы соединяются через фосфатную группу по типу голова-хвост (3′ к 5′) с комплементарными азотистыми основаниями внутри спирали. Такая ориентация цепей называется антипараллельной.
Биосинтез ДНК: реакция в пробирке
Биологов хлебом не корми, а дай воспроизвести биологический процесс в пробирке. Репликация уже давно «приручена» учеными и используется в диагностике.
В 1983 году Кэри Муллис разработал метод, аналогичный биосинтезу ДНК в лаборатории — полимеразную цепную реакцию (ПЦР). В пробирку помещают ферменты, затравки, нуклеозиды и другие необходимые компоненты. Затем запускается цикл биосинтеза интересующих последовательностей ДНК.
Эта схема подходит для генетических тестов:
- выявление предрасположенности к заболеваниям;
- определение индивидуальных особенностей метаболизма;
- установление отцовства;
- поиск преступников.
Трансляция
Трансляция — это синтез полипептидной цепи на матрице иРНК.
Основные органоиды, участвующие в трансляции, — рибосомы. У эукариот рибосомы находятся в митохондриях и пластидах (70S-рибосомы), в свободной форме в цитоплазме (80S-рибосомы) и на мембранах эндоплазматической сети (80S-рибосомы). Синтез белков может происходить в цитоплазме, на шероховатой эндоплазматической сети, в митохондриях и пластидах. В цитоплазме синтезируются белки для нужд клетки, а белки, синтезируемые на ЭПС, транспортируются в комплекс Гольджи и выводятся из клетки. Рибосома состоит из малой и большой субъединиц. Малая субъединица отвечает за декодирование, а большая — за ферментативные функции.
В малой субъединице рибосомы находится функциональный центр (ФЦР) с двумя участками: пептидильным (Р-участок) и аминоацильным (А-участок). В ФЦР могут находиться шесть нуклеотидов иРНК: три в пептидильном и три в аминоацильном участках.
Для транспорта аминокислот к рибосомам используются транспортные РНК (тРНК). Длина тРНК составляет от 75 до 95 нуклеотидов, и они имеют третичную структуру, напоминающую лист клевера. В тРНК различают антикодоновую петлю и акцепторный участок. Антикодон комплементарен кодовому триплету определенной аминокислоты, а акцепторный участок на 3′-конце присоединяет аминокислоту с помощью фермента аминоацил-тРНК-синтетазы (с затратой АТФ). Каждая аминокислота имеет свои тРНК и ферменты, присоединяющие аминокислоту к тРНК.
Двадцать видов аминокислот кодируются 61 кодоном, что теоретически предполагает 61 вид тРНК с соответствующими антикодонами. Однако кодируемых аминокислот всего 20, поэтому у одной аминокислоты может быть несколько тРНК. Установлено, что несколько тРНК могут связываться с одним и тем же кодоном, так как последний нуклеотид в антикодоне не всегда важен. В клетке обнаружено около 40 различных тРНК.
Синтез белка начинается с присоединения малой субъединицы рибосомы к 5′-концу иРНК, в Р-участок которой заходит метиониновая тРНК. Любая полипептидная цепь на N-конце начинается с метионина, который часто отщепляется в дальнейшем. Синтез полипептида происходит от N-конца к С-концу, образуя пептидную связь между карбоксильной группой первой и аминогруппой второй аминокислоты.
Затем присоединяется большая субъединица рибосомы, и во А-участок поступает вторая тРНК, чей антикодон комплементарен кодону иРНК. Пептидилтрансферазный центр большой субъединицы катализирует образование пептидной связи между метионином и второй аминокислотой. Энергия для этого процесса поступает за счет гидролиза ГТФ.
После образования пептидной связи метиониновая тРНК отсоединяется, а рибосома перемещается к следующему кодовому триплету иРНК, который попадает в А-участок. Метиониновая тРНК выталкивается в цитоплазму. На один цикл расходуется 2 молекулы ГТФ. В А-участок заходит третья тРНК, и образуется пептидная связь между второй и третьей аминокислотами.
Трансляция продолжается до тех пор, пока в А-участок не попадает кодон-терминатор (УАА, УАГ или УГА), с которым связывается белковый фактор освобождения. Полипептидная цепь отделяется от тРНК и покидает рибосому, после чего происходит диссоциация субъединиц рибосомы.
Скорость передвижения рибосомы по иРНК составляет 5–6 триплетов в секунду. На синтез белковой молекулы, состоящей из сотен аминокислот, клетке требуется несколько минут. Первым искусственно синтезированным белком был инсулин, состоящий из 51 аминокислотного остатка. Для его получения потребовалось провести 5000 операций, в которых участвовали 10 человек в течение трех лет.
В трансляции выделяют три стадии: а) инициация (образование инициаторного комплекса), б) элонгация (соединение аминокислот), в) терминация (образование терминирующего комплекса).
Биосинтез белков
Биосинтез белков — ключевой процесс анаболизма. Все признаки, свойства и функции клеток и организмов определяются белками. Они недолговечны, и их существование ограничено. В каждой клетке постоянно синтезируются тысячи различных белковых молекул. В начале 50-х годов XX века Ф. Крик сформулировал центральную догму молекулярной биологии: ДНК → РНК → белок. Эта догма утверждает, что способность клетки синтезировать определенные белки наследуется, а информация о последовательности аминокислот в белковой молекуле закодирована в последовательности нуклеотидов ДНК. Участок ДНК, содержащий информацию о первичной структуре конкретного белка, называется геном. Гены не только хранят информацию о последовательности аминокислот в полипептидной цепочке, но и кодируют некоторые виды РНК: рРНК, входящие в состав рибосом, и тРНК, отвечающие за транспорт аминокислот. Биосинтез белка включает два основных этапа: транскрипцию — синтез РНК на матрице ДНК (гена) — и трансляцию — синтез полипептидной цепи.
Реакции матричного синтеза
Это категория химических реакций, происходящих в клетках живых организмов. В ходе этих реакций синтезируются полимерные молекулы по образцу, заложенному в других полимерных молекулах-матрицах. На одной матрице может быть синтезировано неограниченное количество копий. К этой категории относятся репликация, транскрипция, трансляция и обратная транскрипция.
| Название реакции матричного синтеза | Характеристика процесса | Основные компоненты |
|---|---|---|
| Репликация | Синтез ДНК на матрице ДНК | Дезоксирибонуклеозидтрифосфаты, ферменты |
| Транскрипция | Синтез РНК на матрице ДНК | Участок ДНК, рибонуклеозидтрифосфаты, ферменты |
| Трансляция | Синтез полипептида на матрице РНК | Рибосомы, иРНК, аминокислоты, тРНК, АТФ, ГТФ, ферменты |
| Обратная транскрипция | Синтез ДНК на матрице РНК | Дезоксирибонуклеозидтрифосфаты, ферменты |
ДНК (дезоксирибонуклеиновая кислота)
ДНК (дезоксирибонуклеиновая кислота) — это чертеж жизни, содержащий наследственную информацию. Эта макромолекула хранит и передает генетическую информацию из поколения в поколение.
ДНК определяет наследственность и изменчивость живых организмов. Закодированная в ней информация задает программу их развития. Генетические факторы предопределяют жизненный путь человека.
ДНК — одна из трех основных макромолекул (наряду с РНК и белками), обеспечивающая хранение, передачу и реализацию генетической программы. Она содержит информацию о структуре различных видов РНК и белков.
В клетках эукариотов (животных, растений и грибов) ДНК находится в ядре в составе хромосом и в некоторых органоидах (митохондриях и пластидах). В прокариотах (бактериях и археях) кольцевая или линейная молекула ДНК, называемая нуклеоидом, прикреплена к клеточной мембране. У них и у низших эукариотов (например, дрожжей) также встречаются небольшие автономные молекулы ДНК, называемые плазмидами.
ДНК — длинная полимерная молекула, состоящая из повторяющихся нуклеотидов. Каждый нуклеотид включает азотистое основание, сахар (дезоксирибозу) и фосфатную группу. Связи между нуклеотидами образуются за счет дезоксирибозы и фосфатной группы (фосфодиэфирные связи).
В большинстве случаев (за исключением некоторых вирусов с одноцепочечной ДНК) ДНК состоит из двух цепей. Вторичная структура ДНК представляет собой двойную спираль, образованную двумя параллельными неразветвленными полинуклеотидными цепями, закрученными в противоположные стороны.
Пуриновые и пиримидиновые основания расположены внутри спирали, а остатки фосфата и дезоксирибозы — снаружи. Две спирали удерживаются вместе водородными связями между парами азотистых оснований. Тимин (Т) образует водородные связи только с аденином (А), а цитозин (Ц) — только с гуанином (Г). В первой паре оснований две водородные связи, во второй — три.
Эти пары оснований называются комплементарными. Комплементарность — это пространственное соответствие молекул, способствующее образованию водородных связей. Она определяет спиралевидную модель ДНК.
Две спирали в молекуле ДНК комплементарны друг другу. Последовательность нуклеотидов в одной спирали определяет последовательность в другой. В каждой паре оснований одно основание — пуриновое, другое — пиримидиновое. Общее число остатков пуриновых оснований равно числу остатков пиримидиновых.
Таким образом,
- ТИМИН (Т) комплементарен АДЕНИНУ (А),
- ЦИТОЗИН (Ц) комплементарен ГУАНИНУ (Г).
Комплементарность полинуклеотидных цепей является химической основой главной функции ДНК — хранения и передачи наследственных признаков.
Репликация ДНК
Двухспиральная структура ДНК с комплементарными полинуклеотидными цепями позволяет ей самоудваиваться (реплицироваться).
Перед удвоением водородные связи разрываются, и цепи раскручиваются и расходятся. Каждая цепь служит матрицей для синтеза комплементарной цепи.
После разделения цепей происходит саморепликация, в результате которой образуется новая двойная спираль, идентичная исходной.
В итоге формируются две дочерние молекулы ДНК: одна цепь берется из родительской ДНК, а другая синтезируется заново.
Таким образом, структура ДНК сохраняется и передается новому поколению.
Длина полинуклеотидных цепей ДНК практически неограничена. Число пар оснований в двойной спирали варьируется от нескольких тысяч у простейших вирусов до сотен миллионов у человека.
Видеофильм «ДНК. Код Жизни»
Рубрики: Нуклеиновые кислоты
Механизм транскрипции
Ключевым ферментом транскрипции является РНК-полимераза, которая различается по строению у прокариот и эукариот. Однако механизм ее действия остается одинаковым: она наращивает цепь комплементарных рибонуклеотидов, образуя фосфодиэфирные связи между ними.
В качестве матрицы для этого процесса используется ДНК. На ее основе могут синтезироваться различные типы РНК, не только информационные, которые участвуют в белковом синтезе.
Участок матрицы, с которого происходит синтез РНК, называется транскриптоном. Он включает промотор (место для присоединения РНК-полимеразы) и терминатор, на котором синтез останавливается.
Сущность генетического кода
ДНК — это информационная матрица, в которой хранится информация о белках, необходимых организму для роста и жизнедеятельности. Белки состоят из аминокислот, а ДНК (и РНК) — из нуклеотидов. Каждой нуклеотидной последовательности молекулы ДНК соответствует определенная последовательность аминокислот в белках.
В клетке существует 20 видов канонических аминокислот и 4 вида нуклеотидов в ДНК. Каждая аминокислота кодируется триплетом нуклеотидов, ключевыми компонентами которого являются азотистые основания. Этот принцип соответствия называется генетическим кодом, а триплеты оснований — кодонами. Ген — это последовательность кодонов, которая содержит информацию о белке, а также служебные сочетания оснований, такие как старт-кодон и стоп-кодон.
Участие ДНК в биосинтезе белка
ДНК образует устойчивую структуру, в которой ключевым элементом является комплементарное соединение азотистых оснований. Двойная спираль ДНК обеспечивает полное воспроизведение молекулы и считывание отдельных участков при синтезе белков. Этот процесс называется транскрипцией.
Во время транскрипции участок ДНК с определенным геном расплетается, и на одной из цепочек — матричной — синтезируется молекула РНК, копирующая вторую цепочку, называемую кодирующей. Синтез основан на способности оснований образовывать комплементарные пары. В этом процессе участвуют некодирующие области ДНК и фермент РНК-полимераза. РНК служит матрицей для синтеза белка, и в дальнейшем ДНК не участвует.
Окончательная компактизация ДНК
В ядре форма макромолекулы дезоксирибонуклеиновой кислоты (ДНК) становится сложной и компактной в несколько этапов.
- Сначала нить сворачивается в структуру типа соленоида — хроматиновую фибриллу толщиной 30 нм. На этом уровне ДНК сокращает свою длину в 6-10 раз.
- Затем фибрилла, используя специфические скэффолд-белки, образует зигзагообразные петли, что уменьшает линейный размер ДНК в 20-30 раз.
- На следующем этапе формируются плотно упакованные петельные домены, имеющие форму, условно названную «ламповая щетка». Они прикрепляются к внутриядерному белковому матриксу. Толщина таких структур составляет 700 нм, при этом ДНК укорачивается примерно в 200 раз.
- Последний уровень организации — хромосомный. Петельные домены уплотняются настолько, что общее укорочение достигает 10 000 раз. Если длина растянутой молекулы составляет около 5 см, то после упаковки в хромосомы она уменьшается до 5 мкм.
Наивысшего уровня упрощения форма ДНК достигает в метафазе митоза. В этот момент она принимает характерный вид — две хроматиды, соединенные центромерой, которая обеспечивает их расхождение в процессе деления. Интерфазная ДНК организована до доменного уровня и распределяется в ядре клетки без особого порядка. Таким образом, морфология ДНК связана с различными фазами ее существования и отражает особенности функционирования этой важной молекулы.